研究テーマは、「ヒト疾患とタイト結合;隙間の細胞生物学」。
ヒトの体には、血液から隔絶されたいくつかの区域が内包されています。これらの区域は、一層の細胞に覆われることによって、独立した内部環境を保っています。
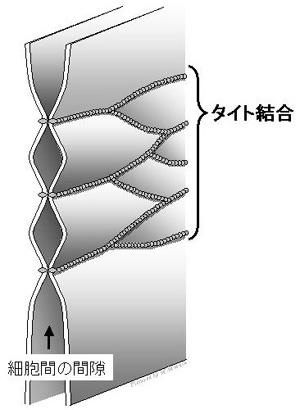
この内部環境保持のために、区域を隔絶している細胞と細胞の隙間を物質が自由に通過できないように厳密にシールしている構造がタイト結合です。もしタイト結合機能が失われると、浮腫、下痢、黄疸のような病態が発症します。
我々の教室では、タイト結合の機能調節機構を分子レベルからヒト疾患のレベルまで含めて解明しようとしています。
タイト結合は、体表を覆う上皮細胞や血管内腔を覆う内皮細胞の細胞間の最も頂部側(すなわち最も外側)に存在する細胞間接着装置です。透過型電子顕微鏡では、隣接する細胞の細胞膜が融合しているように見えることから、密着結合(occludens junction)とも呼ばれています(下図黄矢印)。
| 透過電顕写真(マウス肝臓) |
|---|
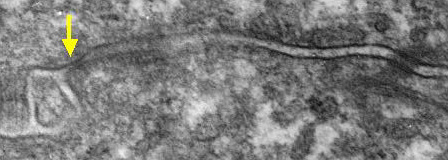 |
| 黄矢印の細胞膜が接している領域がタイト結合。 左側が毛細胆管、右側が血管。 |
凍結割断法で観察すると、タイト結合は、膜内粒子の連続した配列からなるストランドの集合であり、ストランドの数は、タイト結合の機能と関係が深いと考えられています。1998年、この膜内粒子の正体がclaudinという膜貫通型蛋白であることがあきらかになりました。
| 凍結割段写真(マウス肝臓) | タイト結合ストランド |
|---|---|
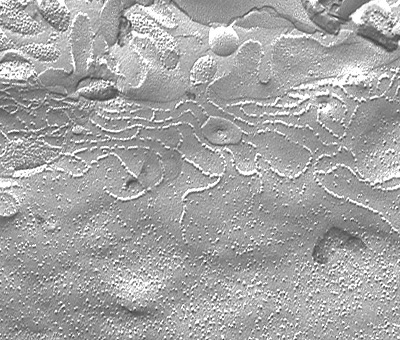
中央部の網状構造物がタイト結合。上部が毛細胆管側。 |
 |
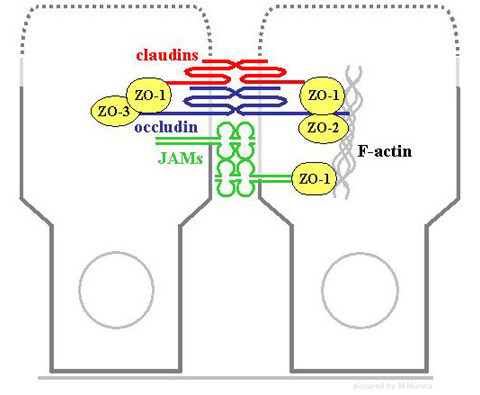
タイト結合は、claudinやoccludin、JAM等の膜貫通型蛋白および、細胞質内の膜裏打ち蛋白という複数の蛋白からなる複合体です。
| 免疫染色写真(マウス肝臓) |
|---|
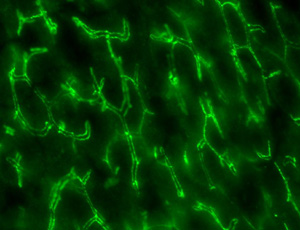 |
| 毛細胆管周囲のclaudinの染色像(緑色) |
タイト結合ストランドを形成できるのは1998年に発見された分子量約23kDaのclaudin のみで、少なくとも20種以上の遺伝子ファミリーからなり、各々が単独でストランドを形成することができます。1個の細胞には通常、複数種のclaudinが発現しており、細胞や臓器の特異性に関与していると考えられています。
occludinは1993年に発見された分子量約65kDaの蛋白で、タイト結合に最も普遍的に存在する蛋白です。
JAMはイムノグロブリンスーパーファミリーに属し、3種の遺伝子ファミリーからなります。
細胞質蛋白は、PDZドメインを含むものと含まないものの2つに大きく分けられます。
前者には、ZO-1、-2、-3、par-3、-6、MAGI-1、-2、-3などがあり、とくにZO-1は膜蛋白を固定する足場蛋白と考えられています。後者には、cingulin、heteromeric G proteins、aPKC、rab-3b、-13、PTENなどが含まれます。
| フェンス機能(ヒト肺腺癌細胞株) |
|---|
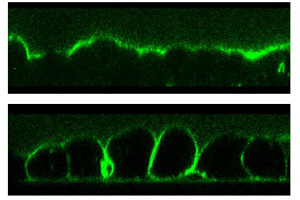 |
| apical側の細胞膜に偏在していた色素(上)がタイト結合の破綻によってbaso-lateral側の細胞膜にも陽性像が見られるようになる(下)。 |
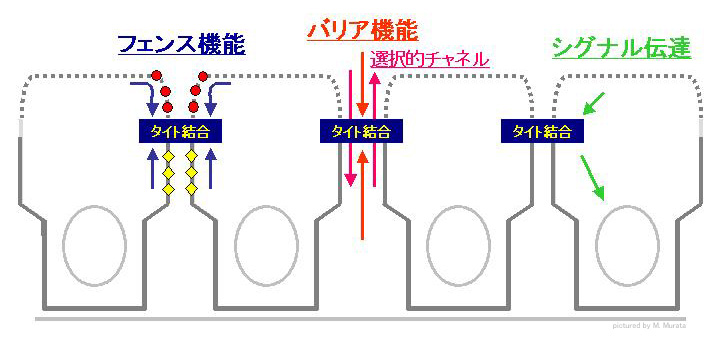
タイト結合の機能のひとつは、細胞膜を区域化し、それを維持することです。この機能はフェンス機能(fence function)と呼ばれており、この機能により、細胞膜は大きく細胞頂部(apical)細胞膜と側壁基底(baso-lateral)細胞膜が分けられており、それぞれの細胞膜に固有の蛋白や脂質の成分は交じり合わないようになっています(左図参照)。言い換えると、細胞の極性を維持する機能です。形態学的に、がん細胞が細胞極性を失い脱分化するのは、タイト結合の形成が低下するためと考えられています。
もうひとつの機能は、細胞と細胞の間(paracellular space)を物質が自由に通過出来ないように細胞間をシールする機能で、バリア機能(barrier function)と呼ばれています。例えば、消化管腔の内外を隔絶する、循環系から胆汁排泄路を隔絶する(下図参照)、血管の内外を分けるといった、区域を隔絶しているのが上皮細胞や内皮細胞間に存在するタイト結合です。
以前は単に物質の通過を阻害していると考えられていましたが、最近では選択的に物質を透過させることができるチャネルとしての役割があることが知られています。
さらに、最近では細胞内への様々なシグナル伝達に関与しているとも考えられています。
ヒトの身体には、半ば独立した内部環境を維持するいくつかの領域を持っています。これらの領域内部の恒常性は、多くの場合、血管内皮細胞が血液とその領域との物質交換を選択的に行うことによって保たれており、このような仕組みを血液臓器関門といいます。
個体の生死を左右する脳浮腫や黄疸などは、血液臓器関門の機能低下症であり、この血液臓器関門を構成する主体が細胞と細胞の隙間をシールしているタイト結合です。浮腫、黄疸は、それぞれ血管内皮、肝細胞のタイト結合機能の異常により起こると考えられています。
| BBBの模式図 |
|---|
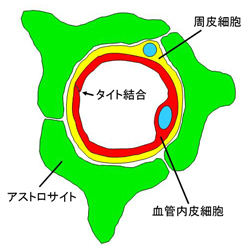 |
血液臓器関門の代表に血液脳関門(blood-brain-barrier: BBB)があります。BBBの本体は、血管内皮細胞に存在するよく発達したタイト結合であり、細胞間隙の物質の通過を厳しく制限して脳内の恒常性維持に深く関与しています。
BBBの機能低下によって生じる最も顕著な所見は、脳浮腫です。脳浮腫とは機能低下したBBB、すなわち血管透過性が亢進した毛細血管から水分が脳の細胞外液腔に過剰に流入した状態を言います。頭蓋内は閉鎖された空間であるため、重篤な脳浮腫により頭蓋内圧が高度に亢進すると、頭痛、嘔吐などの頭蓋内圧亢進症状が出現するようになり、さらに高度なものは脳ヘルニアを起こし死に至ることがあります。このような重篤な脳浮腫をひき起こす疾患として、急速に増大する悪性脳腫瘍、脳膿瘍、脳内血腫、脳塞栓症等の急速な循環障害、また重症頭部外傷など直接血管壁を損傷する場合があります。このような疾患では、病変そのものに対する治療だけではなく、脳浮腫に対する対策が患者の予後を大きく左右することになります。
多くの場合、BBBの破綻は血管透過性の亢進を意昧しており、VEGF(vascular endothelial growth factor)やサイトカインがタイト結合機能を低下させることによって引き起こされます。BBBの調節機構がより詳細に解明され、脳浮腫の効果的な治療法が確立することが期待されています。
肝臓は、胆汁を排泄する外分泌と、アルブミンなどの血清蛋白を血中に放出する広義の内分泌機能を行っています。肝細胞の細胞膜は、毛細胆管の辺縁に存在するタイト結合のフェンス機能によって、毛細胆管膜とそれ以外の細胞膜とに区別されています。
| 透過電顕写真(マウス肝臓) |
|---|
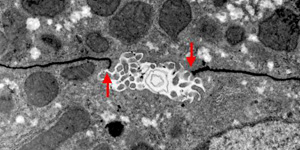 |
| 門脈より注入した色素が赤矢印の部分(タイト結合)で途絶している。 |
肝臓の内分泌機能は類洞という特殊な構造で効率よく営まれ、血液はグリソン鞘から中心静脈へと流れています。一方、外分泌機能、つまり胆汁は肝細胞間に存在する毛細胆管に排泄されます。さらにギャップ結合による細胞間コミュニケーションによって、隣り合う肝細胞が毛細胆管を協調的かつ規則的に収縮させることにより、胆汁は中心静脈側からグリソン鞘側へと送り出され、小葉間胆管に至ります。この二つの相反する流れは、毛細胆管をシールするように存在するタイト結合によってのみ隔てられており、細胞傷害性を持つ胆汁酸は、通常毛細胆管から漏れ出ることはありません。我々はこの肝細胞のタイト結合を血液胆汁関門(blood-biliary-barrier)と呼んでいます。
黄疸、特に直接ビリルビン優位の黄疸は、血液胆汁関門、すなわちタイト結合が機能不全に陥った状態といえます。この原因には、胆石や胆道癌などにより胆管が物理的に閉塞された場合と、胆汁排泄路の閉塞が認められないのに、抱合型ビリルビンが血中に増える場合があり、後者は敗血症等の重症感染症や消化管手術後に認められることがあります。これは腸内から血中に移行したエンドトキシンによって直接的に、またはエンドトキシンによって肝のクッパー細胞から放出されるTNFαなどのサイトカインによって、肝細胞のタイト結合の機能が低下するために起こります。
また、肝内胆汁うっ滞は、肝細胞のタイト結合のみならず、ギャップ結合、微小管とアクチンのいずれか一つの機能が低下しても起こりうることが明らかになってきました。さらに、これらの機能は互いに調節し合っており、肝細胞のタイト結合に直接作用する薬物だけでなく、ギャップ結合やアクチンの機能に影響を与える薬物も、タイト結合のバリア機能を間接的に低下させ、肝内胆汁うっ滞や直接ビリルビン優位の黄疸をひき起こす可能性があると考えられています。
網膜の血液循環には網膜循環と脈絡膜循環があり、それぞれがバリアを有しており、その両者を併せて血液網膜関門blood retinal barrier(以下BRB)を形成し網膜のホメオスタシスを維持しています。網膜循環では毛細血管内皮細胞が内側血液網膜関門inner BRB(以下i-BRB)を形成しますが、脈絡膜系の血管は透過性が高いため網膜色素上皮細胞が外側血液網膜関門となって、血液成分が感覚網膜に侵入するのを制限しています。特にi-BRBは、網膜内の恒常性維持に重要な役割を果たしているので、その破綻は、黄斑浮腫を代表とする病変を引き起こし、糖尿病網膜症患者の視力低下や失明を招きます。糖尿病網膜症は、わが国における後天性失明の原因の第一位で、毎年約4,000人がこの病気で失明しているといわれています。
糖尿病網膜症では、大まかに網膜毛細血管の透過性亢進から内腔閉塞、血管新生へと進行し、最終的に失明へと至ります。したがって、糖尿病網膜症の初期変化である網膜毛細血管の透過性亢進を防ぐことができれば、その進行をくい止めることができる可能性があります。糖尿病において網膜毛細血管の透過性を亢進させる引き金は、網膜でVEGFが産生されるようになるためであることがわかってきました。VEGFは血管の透過性を亢進させる因子であり、現在ではタイト結合の構成蛋白質をリン酸化し、タイト結合バリア機能を低下させることが明らかになっています。VEGFを産生する細胞は、普段はGDNF (glial-cell-line-derived neurotrophic factor)を産生して血管透過性を下げているミューラー細胞であり、ミューラー細胞の性質を正常状態に保つ、あるいは糖尿病状態から正常に戻すことができれば、糖尿病網膜症による失明を防ぐことができると考えられています。
これら以外にも血液胸腺関門と免疫の関係、最近話題となっている環境ホルモンの作用と血液精巣関門の問題、癌の血行転移と血管のタイト結合など、血液臓器関門は様々な病態の理解に重要なものとなっています。
さらに2002年には、体の表面を覆う表皮にもタイト結合が存在し、体外への水分の漏出を防いでいることが示され、外界から生体を隔離する重要な装置であることが明らかになりました。
また、Claudin-14は内耳の外有毛細胞に、claudin-16/paracellin-1は腎のHenle上行脚に発現しそれらの遺伝子の変異によって、それぞれ遺伝性難聴、遺伝性低マグネシウム血症を引き起こすことが知られています。
一部のタイト結合蛋白(claudin-3、-4、JAM)は、細菌毒素およびウイルスのレセプターとなるとも考えられています。
このようにタイト結合が様々な病態に関与していることが明らかになりつつありますが、タイト結合の構成分子が明らかになったのは1998年であり、今後、タイト結合そのものの調節機構が明らかになるにつれて、血液臓器関門の調節機構と疾患の関係がますますはっきりとしてくると考えられています。