丂巹偨偪偺惗懱婡擻壔妛尋媶幒偼丆慜擟偺忋栰捈恖愭惗偑婎惗尋偵堏傜傟偰偐傜侾擭埲忋宱偭偨嶐擭弔偵丆慡偔怴偟偄恮梕偱僗僼傿儞僑帀幙傗摐帀幙偺惗懱婡擻偺夝柧傪傔偞偟偰嵞弌敪偟偨偽偐傝偺怴偟偄儔儃偱偡丏巹偼婣崙偡傞傑偱侾俈擭娫奜崙偱尋媶惗妶傪憲偭偰偒傑偟偨偑丆摿偵嵟嬤侾侽擭偼僔傾僩儖偵偁傞儚僔儞僩儞戝妛僶僀僆儊儞僽儔儞尋媶強乮敔庣愬堦榊強挿乯傗僼儗僢僪僴僢僠儞僜儞娻尋媶強偱僗僼傿儞僑帀幙偺惗棟婡擻偵娭偡傞尋媶傪恑傔偰偒傑偟偨丏偙傟傑偱斾妑揑抧枴側嵶朎枌帀幙偺堦偮偲巚傢傟偰偄偨僗僼傿儞僑帀幙傕偙偺侾侽擭偺尋媶偱丆偦偺暘夝嶻暔偱偁傞僙儔儈僪傗僗僼傿儞僑僔儞傗偦偺桿摫懱偺僔僌僫儖揱払偵偍偗傞栶妱偑拲栚偝傟傞傛偆偵側傝傑偟偨丏峏偵嬌偔嵟嬤偱偼丆僗僼傿儞僑帀幙傗摐帀幙偼丆怴偨側嵶朎枌峔憿奣擮偲偟偰拲栚傪庴偗偰偄傞僨僞乕僕僃儞僩晄梟惈偺儈僋儘僪儊僀儞峔憿偺拞怱揑側峔惉惉暘偲偟偰丆偦偙偵偍偗傞僔僌僫儖暘巕偺暘攝傗桝憲偵偍偗傞廳梫側栶妱傪偵側偭偰偄傞偙偲偑帵嵈偝傟丆儂僢僩側尋媶椞堟偵側傝偮偮偁傝傑偡丏丂巹偨偪偼偦偙偱丆傾儊儕僇偱偺尋媶傪擔杮偱峏偵怴偟偄儗儀儖偵敪揥偝偣傞傋偔丆怴偟偔彆嫵庼偵摐帀幙傪偙傟傑偱尋媶偟偰偒偨堜僲岥恗堦丆彆庤偵庒偄暘巕惗暔妛幰偱偁傞杮嫶寬丆榓揷弤傪寎偊偰尋媶傪僗僞乕僩偝偣傑偟偨丏偙傟偵戝妛堾惗俈柤丆懖尋惗俇柤側偳傪壛偊偰憤惃侾俉柤偺恮梕偱婃挘偭偰偍傝傑偡丏
丂偙偆偟偨條乆側尋媶偺側偐偱巹偨偪偑崱堦斣娭怱傪帩偭偰廤拞揑偵尋媶傪偟偰偄傞偺偼僗僼傿儞僑僔儞侾亅儕儞巁偺惗棟婡擻偵娭偟偰偱偡丏恾偵帵偟偨傛偆偵丆僗僼傿儞僑僔儞侾亅儕儞巁偼堦偮偺暘巕偱偁傝側偑傜嵶朎撪僙僇儞僪儊僢僙儞僕儍乕偲偟偰偺栶妱偲丆嵶朎偺奜偐傜摥偔僙僇儞僪傾僑僯僗僩偲偟偰偺栶妱偑摨帪偵峫偊傜傟偰偄傞偲偰傕儐僯乕僋側帀幙儊僨傿僄乕僞乕偱偁傝傑偡丏1991擭偵丆巹偼摉帪恑傔偰偄偨僗僼傿儞僑僔儞桿摫懱偺娻揮堏梷惂偺尋媶偺夁掱偱丆偦傟傑偱堦斒偵偼丆僗僼傿儞僑僔儞偺戙幱拞娫懱偵夁偓側偄偲傒側偝傟偰偄偨僗僼傿儞僑僔儞侾亅儕儞巁偑儃僀僨儞僠儍儞僶乕朄側偳偵傛傞専嶕偱丆嬼慠儊儔僲乕儅側偳偁傞庬偺娻嵶朎偺憱壔惈塣摦傪嬌傔偰掅擹搙偱梷惂偡傞帠幚傪尒偮偗偨偺偑杮尋媶偺奐巒偝傟偨偒偭偐偗偱偟偨乮PNAS, 1992乯丏偦傟偼僕儑乕僕僞僂儞戝妛偺Spiegel傜偑嵶朎憹怋偺嵶朎撪儊僢僙儞僕儍乕偲偟偰偺栶妱傪敪昞偟偨偺偲傎傏摨偠偙傠偱偟偨丏巹偨偪偼峏偵丆儚僔儞僩儞戝妛偺Ross嫵庼傜偲偺嫟摨尋媶偱丆僸僩暯妸嬝嵶朎偼偙偺帀幙偵嬌傔偰姶庴惈偑崅偔丆PDGF埶懚惈偺憱壔惈塣摦偵懳偟偰傕梷惂嶌梡傪傕偮偙偲傪尒偄偩偟丆偦偺儊僇僯僘儉偺尋媶傪恑傔偰偒傑偟偨 (JBC, 1995乯丏峏偵寣彫斅偱偼暘壔偺夁掱偱僗僼傿儞僑僔儞侾亅儕儞巁暘夝峺慺偑寚懝偟偦偺偨傔偙偺帀幙偑寣彫斅偵拁愊偡傞帠幚偵拝栚偟丆寣彫斅偵偍偗傞尋媶傕摨帪偵奐巒偟丆妶惈壔偝傟偨寣彫斅偐傜僗僼傿儞僑僔儞侾亅儕儞巁偑曻弌偝傟丆偙傟偑嵶朎偺奜偐傜僆乕僞僐僀僪偲偟偰寣彫斅妶惈壔傪槾恑偟偰偄傞帠幚傪撍偒偲傔傑偟偨乮Blood, 1995乯丏偦偟偰偙偺帀幙傪晄摦壔偟偨崌惉價乕僘傗曻幩儔儀儖帀幙崌惉傪摫擖偡傞偙偲偵傛傝丆僗僼傿儞僑僔儞侾亅儕儞巁偑嵶朎枌偵懚嵼偡傞庴梕懱傪夘偟偰嶌梡偡傞偲偄偆奣擮傪偄偪憗偔採彞偟乮JBC, 1997乯丆嵶朎枌忋偺寢崌抈敀偺摨掕傪帋傒偰偒傑偟偨乮Biochemistry, 1998乯丏偦偟偰嶐擭偵側偭偰幚嵺偵偙偺帀幙偺庴梕懱堚揱巕偑偄偔偮偐偺儔儃偱摨掕偝傟傞偵媦傫偱丆LPA偲摨條側嵶朎娫儊僨傿僄乕僞乕偲偟偰偺栶妱偑傎傏妋棫偟丆偁傜偨傔偰惗棟揑偁傞偄偼昦棟揑忦審壓偵偍偗傞偙偺惗棟妶惈帀幙偺摦懺偑峀偔拲栚偝傟傞偵帄偭偰偄傑偡丏丂偙傟偵娭偟偰偼巹偨偪偺尋媶幒偱偼尰嵼壓偺傛偆側僾儘僕僃僋僩偑恑峴偟偰偄傑偡丏
1丏 僗僼傿儞僑僔儞亅侾亅儕儞巁庴梕懱偺婡擻夝愅
丂嵟嬤変乆偼偙偺1擭娫偺尋媶偱儅僂僗偵敪尰偟偰偄傞僗僼傿儞僑僔儞亅侾亅儕儞巁偺庴梕懱偲偟偰丆偙傟傑偱偵抦傜傟偰偄偨Edg1, Edg2偺奜偵怴偨偵Edg3, Edg5丆偝傜偵偼偙傟傜偲憡摨惈傪帩偮Edg6側偳偺庴梕懱抈敀傪僋儘乕僯儞僌偟傑偟偨丏偦偙偱尰嵼偙偆偟偨庴梕懱偺婡擻傪偦傟偧傟偺嫮惂敪尰姅傪嶌惢偟偨傝偟側偑傜夝愅傪恑傔偰偄傑偡丏傑偨偦偺僔僌僫儖揱払婡峔傪扵傞栚揑偱Rho, Rac, cdc42側偳G抈敀幙偺妶惈壔宆丆桪惃晄擻曄堎懱傪儊儔僲乕儅嵶朎丆慄堐夎嵶朎偵嫮惂敪尰丆傑偨偼旝検拲擖傪偟偨傝丆峺曣僣乕僴僀僽儕僢僪朄偵傛偭偰庴梕懱偲楢娭偟偰偄傞暘巕傪専嶕偟偨傝偟偰丆偙偺帀幙偺帵偡惗暔妶惈偺嵶朎摿堎惈偺暘巕婡峔忋偺堄枴晅偗傪傔偞偟傑偡丏
2丏 僗僼傿儞僑僔儞侾亅儕儞巁偺嵶朎偐傜偺曻弌婡峔
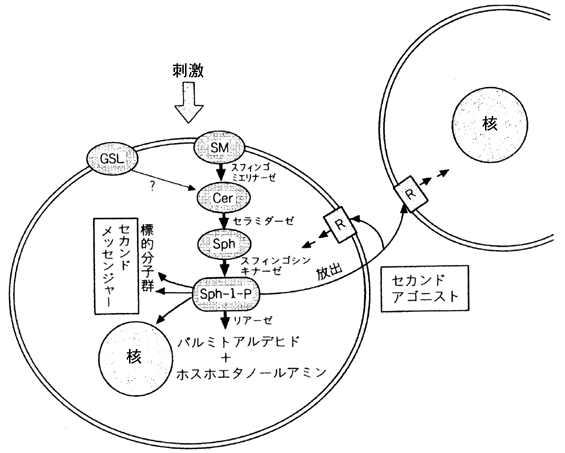
丂妶惈壔偝傟偨寣彫斅偐傜偺僗僼傿儞僑僔儞侾亅儕儞巁偺曻弌婡峔偼偦偺PKC埶懚惈埲奜傎偲傫偳晄柧側傑傑偱偡丏偦偙偱丆尰嵼丆庬乆偺慾奞嵻傪巊偭偨惗壔妛揑尋媶偲偲傕偵嫄妀媴嵶朎傪儌僨儖偵曻弌偺暘巕婡峔偺夝柧傪捛媮偟偰偄傑偡丏傑偨寣燋拞偵曻弌偝傟偨僗僼傿儞僑僔儞侾亅儕儞巁偑偳偺傛偆側婡彉偱暘夝徚柵偝傟傞偺偐寣燋拞偵偼偦偺暘夝峺慺偼懚嵼偟側偄偨傔崱偺偲偙傠慡偔晄柧偱丆偙傟傕廳梫側僥乕儅偲側偭偰偄傑偡丏
3丏 僗僼傿儞僑僔儞僉僫乕僛偺懡條惈偲婡擻挷愡
丂僗僼傿儞僑僔儞侾亅儕儞巁偼嵶朎撪偱僗僼傿儞僑僔儞僉僫乕僛偲偄偆峺慺偵傛偭偰崌惉偝傟傑偡丏庬乆偺奜晹巋寖傪庴偗偰妶惈壔偝傟偨杮峺慺偵傛偭偰堦夁惈偵偮偔傜傟偨僗僼傿儞僑僔儞侾亅儕儞巁偑IP3偲偼堎側傞婡峔偱Ca2亄摦堳側偳傪堷偒婲偙偡偲偄偆乽僗僼傿儞僑僔儞僉僫乕僛夞楬乮SK夞楬乯乿偺懚嵼偑儅僗僩嵶朎偺妶惈壔傗慄堐夎嵶朎偺憹怋丆暯妸嬝嵶朎偺塣摦側偳偵偍偄偰廳梫側栶妱傪壥偨偟偰偄傞偲峫偊傜傟偰偄傑偡丏偙偺Key峺慺偺徻偟偄妶惈壔婡峔偼峺慺帺懱偺尋媶偑抶傟偰偄偨偙偲傕偁傝丆傎偲傫偳庤偮偐偢偺忬懺偱偁傝傑偟偨丏偟偐偟嶐擭偵側偭偰Spiegel傜偵傛偭偰偙偺峺慺偺惛惢偲cDNA僋儘乕僯儞僌偑惉偟悑偘傜傟偨偙偲偵傛傝丆傛偆傗偔怴偟偄敪揥偺壜擻惈偑奐偗偰偒偨偲偄偊傑偡丏巹偨偪傕丆偙傟傑偱偺梊旛揑側幚尡偐傜慻怐摿堎揑側杮峺慺偺暘巕懡條惈傪憐掕偟偰偦偺扵嶕傪帋傒偰偍傝傑偡丏傑偨偦偺妶惈挷愡婡峔乮妶惈壔場巕偺摨掕丆慾奞場巕偺晄妶惈壔丆嵶朎撪堏峴偺桳柍側偳乯傪惗壔妛揑丆暘巕惗暔妛揑庤朄偱夝柧偡傞偙偲偵傛偭偰丆SK夞楬偺扴偭偰偄傞惗棟揑栶妱傪柧傜偐偵偟丆峏偵偼傾儗儖僊乕傗摦柆峝壔偵偍偗傞杮峺慺偺桳岠側慾奞嵻偺扵嶕傗SK夞楬偺惂屼場巕傪摨掕偟丆怴偟偄堛栻奐敪偺帋傒偺堦彆偲偟偰偄偒偨偄偲峫偊偰幚尡偵庢傝慻傫偱偄傑偡丏
丂偦傟埲奜偵傕丆杮尋媶幒偱偼
4丏 僗僼傿儞僑帀幙嵶朎撪寢崌僞儞僷僋幙偺扵嶕
5丏 僗僼傿儞僑帀幙丆摐帀幙偺嵶朎枌敵峔憿偵偍偗傞栶妱
6丏 摐帀幙偵傛傞恄宱嵶朎偺暘壔丆婡擻傊偺塭嬁
側偳偺壽戣偵偦傟偧傟偺僌儖乕僾偑庢傝慻傫偱偄傑偡丏巹帺恎偼惗壔妛偑僶僢僋偺尋媶傪偙傟傑偱恑傔偰偒傑偟偨偟丆帀幙偼堚揱忣曬偺捈愙嶻暔偱偼側偄偨傔丆暘巕惗暔妛偺愴棯偐傜庢傝巆偝傟傞偒傜偄偑偁傝傑偟偨偑丆嵟嬤偱偼忋弎偟傑偟偨傛偆偵庴梕懱傗娭楢峺慺偺僋儘乕僯儞僌偑恑傓側偳怴偟偄敪揥傪傒偣偰偄傑偡丏傑偨偦傟偩偗偱側偔丆帀幙偺尋媶偵偼丆僎僲儉尋媶偺榞偵偼廂傑傝偒傜側偄怴偟偄愴棯丆曽朄榑偑昁梫偱偁傞偙偲偼柧傜偐偱丆帀幙亅抈敀幙憡娭側偳偺娤揰偐傜丆偙傟傑偱偺暘巕惗暔妛偵寚偗偰偄傞偲巚傢傟傞柺傪孈傝婲偙偟丆惗暔妛偺怴偟偄敪揥偵峷專偱偒偨傜偲婅偭偰偍傝傑偡丏
俈丏娭楢憤愢暥專
- Hakomori S. & Igarashi Y. Functional role of glycosphingolipids in cell recognition and signaling. J. Biochem. (invited review) 118 : 1091-1103, 1995.
- 屲廫棐桋擵丂僗僼傿儞僑僔儞1-儕儞巁偺惗棟妛亅嵶朎塣摦惂屼婡擻偲寣彫斅偵偍偗傞婡擻揑栶妱傪拞怱偲偟偰亅丂惗壔妛69 (10) : 1166-1185, 1997.
- Igarashi Y. Functional roles of sphingosine, sphingosine 1-phosphate and methylsphingosines : in regards to membrane sphingolipid signaling pathway. (invited review) J. Biochem. 122 (6) 1080-1087, 1997.
- 屲廫棐桋擵丂儊僠儖僗僼傿儞僑僔儞偺惗棟丒栻棟嶌梡亅僗僼傿儞僑帀幙僔僌僫儖揱払偲偺娭學偲帀梟惈栻嵻摫擖偵岦偗偰偺堦偮偺傾僾儘乕僠亅丂抈敀幙丂妀巁丂峺慺丂43 : 226-236, 1998.
- Igarashi Y. Sphingosine 1-phosphate as an intercellular signaling molecule. Ann. N. Y. Acad Sci. "Sphingolipids as singaling modulators in the nervous system" 845 : 19-31, 1998.
- 栴晉桾丆屲廫棐桋擵丆旜嶈桼婎抝丂僗僼傿儞僑僔儞1-儕儞巁偵傛傞忣曬揱払丏抈敀幙丂妀巁丂峺慺丂43 (16) : 2510-2515, 1998.
- 屲廫棐桋擵丂僙僇儞僪儊僢僙儞僕儍乕偁傞偄偼僙僇儞僪傾僑僯僗僩偲偟偰偺僗僼傿儞僑僔儞1-儕儞巁尋媶偺怴偟偄揥奐丏抈敀幙丂妀巁丂峺慺丂44 (7) : 112-119, 1999.