杒奀摴戝妛丒擾妛尋媶壢丒墳梡惗柦壢妛愱峌
丂戞侾侾夞ICV偑侾俋俋俋擭俉寧俋擔偐傜侾俁擔傑偱丆僆乕僗僩儔儕傾偺僔僪僯乕偱奐嵜偝傟偨丏ICV偵偼枅夞嶲壛偟偰偄傞傢偗偱偼側偄偑丆崱夞偼僆乕僗僩儔儕傾偲偄偆怱抧傛偄尵梩偺嬁偒偵桿傢傟偰丆擾妛晹偺怉暔僂僀儖僗尋媶幒偐傜偼嫵姱俁恖慡堳偑偺偙偺偙偲弌偐偗偨傢偗偱偁傞丏擔杮偱偼偍杶媥傒偺恀偭嵟拞偱丆僆乕僗僩儔儕傾墲暅偺旘峴婡傕枮惾忬懺偱偁偭偨丏丂崙嵺妛夛偵嶲壛偡傞偲儔僀僶儖偨偪偑偄偭偨偄偳偙傑偱恑傫偱偄傞偺偐偮偄偮偄婥偵側偭偰偟傑偆丏僾儘僌儔儉傪傕傜偆傗偄側傗億僗僞乕僙僢僔儑儞傗僔儞億僕僂儉偵嵶偐側僠僃僢僋傪偄傟傞偺偼巹偩偗偱偼側偄偩傠偆丏帺暘偺愱栧偱偁傞怉暔僂僀儖僗偺敪昞偼傕偪傠傫摦暔僂僀儖僗偺尋媶敪昞偵傕尒棊偲偟偑側偄偐婷梸偵側偭偰偄傞帺暘傪敪尒偡傞丏嬤擭丆愱栧嶨帍偑抣忋偑傝偟偰峸撉傪傗傔偨傕偺偑偨偔偝傫偁傞偙偲傗僨僕僞儖壔偑恑傫偱崱傑偱傁傜傁傜傔偔偭偰傒偰偄偨榑暥傪僀儞僞乕僱僢僩偺儁乕僕偱尒傞傛偆偵側偭偨偙偲偼丆帺暘偲偼慡偔柍娭學偺暘栰偵嬼慠傾僋僙僗偡傞僠儍儞僗傪扗偭偰偄傞丏崙嵺妛夛偼尋媶幰偨偪偑捈偵媍榑偱偒傞儊儕僢僩偩偗偱偼側偔丆嫃傞偩偗偱暘栰奜偺怴慛側忣曬偵傕怗傟傜傟傞旕忢偵岲搒崌偺応強偱偁傞丏摿偵幚尡媄弍傗婡婍偺忣曬偵偮偄偰偼丆帺暘偺尋媶偵傕巊偊傞偺偱偼偲偢偄傇傫偨偔偝傫偺儊儌傪偲偭偰偒偨乮擔杮偱尒捈偡偲側偤偐堄枴晄柧偺傕偺偑懡偐偭偨乯丏
丂偝偰丆ICV偱昅幰偑廂廤偟偨怉暔僂僀儖僗傗庒姳偺摦暔僂僀儖僗偵娭偡傞尋媶忣曬傪偙偙偱徻嵶偵彂偒楢偹偰傕撉幰偺奆偝傫偵偼戅孅側偩偗偱偁傠偆丏傑偨丆尋媶婡婍側偳偺忣曬偼偳偙偐偺儊乕僇乕偐傜幱嬥偱傕偄偨偩偐側偗傟偽彂偔婥偵側傜側偄丏棷妛懱尡婰偵偁傞傛偆側偍傕偟傠偄僄僺僜乕僪偼傢偢偐侾廡娫庛偺懾嵼拞偵婜懸偱偒傞偼偢傕側偄丏怓乆偲巚偄擸傫偩枛丆崱丆怉暔暘巕惗暔妛偱傕偭偲傕儂僢僩側榖戣偲側偭偰偄傞gene silencing偵偮偄偰ICV偱庤偵擖傟偨忣曬傪傕偲偵徯夘偡傞偙偲偵偡傞丏嵟嬤丆Cell丆Science丆Nature偲偄偭偨嶨帍偵栚敀墴偟偺尋媶僥乕儅偱偁傞丏偙偙偱乽壗偩怉暔偺榖偐乿偲偡偖偵擺摼偟側偄偱偄偨偩偒偨偄丏摦暔偱偺尋媶傛傝傕怉暔偱偺尋媶偑侾曕儕乕僪偟偰偄傞傑偝偵怉暔偺尋媶幰偵偲偭偰偼婲巰夞惗偺榖戣側偺偱偁傞丏怉暔偺尋媶偼偙傟傑偱忢偵摦暔偺尋媶偺僐僺乕丒墳梡偵傛偭偰敪揥偟偰偒偨偲尵偭偰傕夁尵偱偼側偄丏摿偵僔僌僫儖僩儔儞僗僟僋僔儑儞偵娭偡傞尋媶偼姰慡偵屻恛傪攓偟偰偄傞乮怉暔偵儐僯乕僋側傕偺傕偨偔偝傫偁傞偼偢側偺偩偑乯丏偙傟偼怉暔偺尋媶幰偺悢偑摦暔偺尋媶幰傛傝傕埑搢揑偵彮側偄偐傜偩偲庡挘偡傞偺偼晧偗惿偟傒偩傠偆偐丏
丂Gene silencing偼嵶朎偵怤擖偟偰偔傞奜棃堚揱巕偺敪尰傪僔儍僢僩傾僂僩偡傞婡峔偱偁傝丆怉暔嵶朎偱傑偢尒弌偝傟偨丏尰嵼傑偱偵怉暔僂僀儖僗偵懳偡傞堦偮偺杊屼婡峔偲偟偰婡擻偡傞偙偲偑徹柧偝傟偰偄傞乮偟偨偑偭偰怉暔僂僀儖僗妛幰偑尋媶傪儕乕僪偟偰偒偨乯丏嵟嬤偵側偭偰丆慄拵丆嬠椶丆崺拵偦偟偰尨惗摦暔偱傕摨條偺尰徾偑敪尒偝傟傞偵媦傫偱丆惗暔嵶朎慡斒偵懚嵼偡傞僔僗僥儉偲峫偊傜傟傞傛偆偵側偭偰偒偰偄傞丏傎擕椶偺嵶朎偱敪尒偝傟傞擔傕嬤偄偺偱偼側偐傠偆偐丏梋択偩偑丆apoptosis乮programed cell death乯偺尋媶偑摦暔嵶朎偱偄偭偒偵崅梘偟偨帪婜偵丆柶塽僔僗僥儉傗敪惗偑懚嵼偟側偄怉暔偵偼柍墢偺尰徾偲巚傢傟偨丏偟偐偟丆尰嵼傑偱偵hypersensitive reaction偲屇偽傟傞昦尨懱偵懳偡傞怉暔偺斀墳偑apoptosis偵崜帡偟偰偄傞偲偄偆曬崘偑悢懡偔弌偝傟偰偄傞丏gene silencing偺尋媶偑偪傚偆偳apoptosis偲偼斀懳偺弴彉偱恑傫偱偄傞偺偐傕偟傟側偄丏
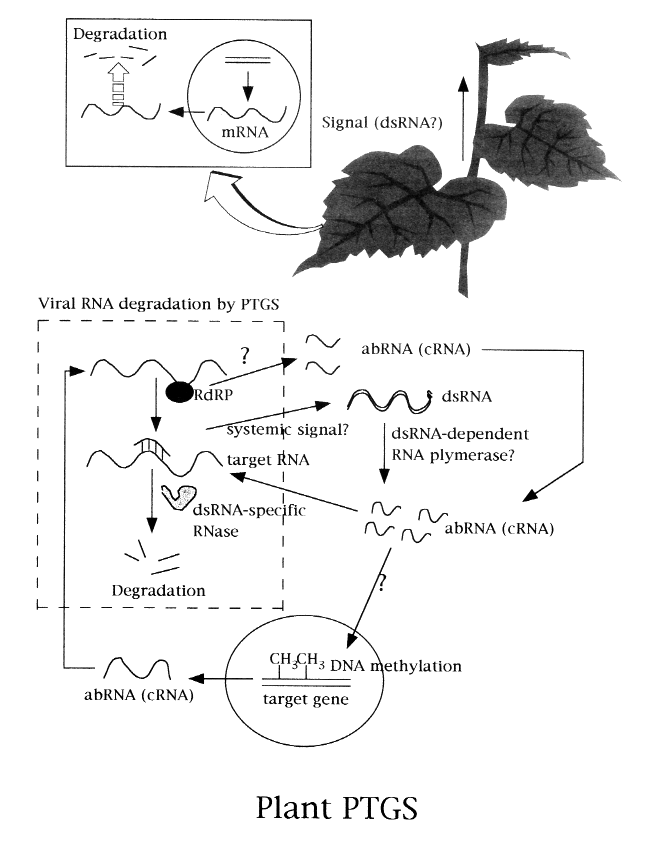
丂Gene silencing偵傕偄偔偮偐偺僞僀僾偑偁傞傛偆偩偑丆偙偙偱偼posttranscriptional gene silencing (PTGS)偵偮偄偰徯夘偡傞乮憓擖恾嶲徠乯丏傕偲傕偲偼丆崅敪尰傪栚巜偟偰嶌弌偟偨宍幙揮姺怉暔偺transgene偺敪尰偑媡偵姰慡偵梷惂偝傟傞偲偄偆尰徾偑娤嶡偝傟丆偙偺尰徾傪gene silencing偲屇傇傛偆偵側偭偨丏偙偺帪丆妀偱偺揮幨偼惓忢偵婲偙傝丆嵶朎幙偵堏摦偟偨屻偵transgene偺mRNA偑暘夝偝傟傞傕偺傪PTGS偲掕媊偟偨丏PTGS傪婲偙偟偰偄傞怉暔嵶朎偱偼transgene偺妀DNA偑摿堎揑偵儊僠儖壔偝傟傞偲曬崘偝傟偰偄傞丏偝偰丆偙偺transgene偑僂僀儖僗偺僎僲儉堚揱巕偱偁傞帪偵偼丆偦偺怉暔懱偼僂僀儖僗掞峈惈乮偁傞偄偼懴惈乯偲側傞丏PTGS桿摫偵偼丆嵶朎幙偱偺RNA擹搙偑偁傞堦掕偺thresh-hold傪墇偊傞昁梫偑偁傞偲峫偊傜傟偰偄傞丏RNA僂僀儖僗偑嵶朎撪偱憹怋偟偨応崌丆偦偺thresh-hold傪墇偊傞偩傠偆偲梊憐偝傟丆偙偺帪偵桿摫偝傟偨PTGS偵傛偭偰丆僂僀儖僗RNA偼暘夝偝傟傞丏偄偭偨偄偳偆偄偆暘巕儗儀儖偱偺儊僇僯僘儉偑摥偄偰偄傞偺偩傠偆偐丏偙偺媈栤偵偮偄偰傕丆傢偢偐偙偙俀丆俁擭偱懡偔偺徹嫆偑摼傜傟偰偄傞丏transgene偐傜偺mRNA偺揮幨検偑懡偄応崌丆壗傜偐偺僔僗僥儉乮RNA埶懚RNA polymerase: RdRP丠乯偵傛偭偰dsRNA偑宍惉偝傟丆偁傞偄偼挿偝偺抁偄堎忢側RNA (abRNA)偑崌惉偝傟丆偙傟傜偑僔僌僫儖偲側偭偰巘晹傪捠傝慡恎偵堏峴偡傞丏僔僌僫儖偼僈僀僪僞儞僷僋傗儗僙僾僞乕側偳偺摿堎揑側僔僗僥儉傪夘偟偰妀偺拞偵堏峴偟丆僞乕僎僢僩DNA傪儊僠儖壔偡傞偲峫偊傜傟偰偄傞丏偙偺儊僠儖壔堚揱巕偐傜偺揮幨嶻暔傕傑abRNA偲側傝丆偙傟偑complementarity偺偁傞RNA偵hybridize偟偰晹暘揑側dsRNA傪宍惉偡傞丏嵟廔揑偵偦偺僞乕僎僢僩RNA偼dsRNA摿堎揑RNase偵傛偭偰暘夝偝傟傞丏偦傟偱偼transgene傪帩偭偰偄側偄怉暔偵僂僀儖僗傪姶愼偝偣偨応崌丆PTGS偼桿摫偝傟傞偺偐丏cross protection偲屇偽傟傞尰徾偑偁傞丏偁傜偐偠傔丆昦尨椡偺庛偄僂僀儖僗傪愙庬偟偰偍偒丆屻偐傜昦尨椡偺嫮偄僂僀儖僗傪愙庬偟偰傕昦婥傪婲偙偝側偄偲偄偆尰徾偱丆摦暔偺柶塽僔僗僥儉偵帡偰偄傞乮尵偆傑偱傕側偔怉暔偵偼柶塽僔僗僥儉偼懚嵼偟側偄乯丏幚偼丆偙偺crossprotection傪PTGS偱愢柧偡傞偙偲偑偱偒傞丏偡側傢偪丆怉暔僂僀儖僗偑姶愼偡傞偲廻庡偺RdRP偺敪尰傪桿摫偡傞丏偙傟偑丆僂僀儖僗RNA傪拻宆偵偟偰懡偔偺abRNA傪崌惉偟丆僂僀儖僗RNA偵寢崌偟偰暘夝偡傞乮偁傞偄偼dsRNA摿堎揑RNase偑暘夝偡傞乯丏偙偺応崌丆妀DNA忋偵偼僂僀儖僗攝楍偼懚嵼偟側偄偺偱僔僌僫儖傪夘偟偨儊僠儖壔偼婲偒側偄偺偱偁傞丏
丂嵟屻偵丆怉暔僂僀儖僗偺尋媶幰偑偙偺PTGS偵娭偟偰幚偵嫽枴怺偄尰徾傪敪尒偟偰偄傞偙偲傪弎傋偰偍偒偨偄丏幚偼怉暔僂僀儖僗偺拞偵偙偺PTGS傪梷惂偡傞偨傔偺堚揱巕乮PTGS suppressor乯傪偁偊偰僐乕僪偟偰偄傞傕偺偑懚嵼偡傞偺偱偁傞丏Potyvirus偺HC-Pro傗cucumber mosaic virus偺2b僞儞僷僋側偳偑偦傟偱偁傞乮HC-Pro偲2b偲偺娫偵偼homology偼側偄乯丏PTGS suppressor偼丆僂僀儖僗偲廻庡偺挿偄峌杊偺楌巎偺拞偱僂僀儖僗偑岻傒偵妉摼偟偨傕偺偱偁傠偆丏
_____________________________
Chikara Masuta
Plant Virology Lab.
Faculty of Agriculture
Hokkaido University
kita-ku, kita 9, nishi 9
Sapporo 060-8589, Japan
Phone & FAX:81-11-706-2483
masuta@res.agr.hokudai.ac.jp
{kind=link}