肺がん
ページ内目次
肺サ—ファクタント蛋白質によるEGFシグナル制御機構の解明
肺がん治療において上皮増殖因子受容体(EGFR)は最も重要な標的分子の一つであり、EGFRチロシンリン酸化阻害薬が実用化され、肺がん治療は大きく進歩したと言えます。しかしながら効果がEGFRのチロシンキナーゼドメインに遺伝子変異を有する症例に限られることや、奏効例もやがて耐性を獲得するといった課題もあるのが現状です。
肺サーファクタント蛋白質AとD(SP-A、SP-D)は肺胞Ⅱ型細胞やクララ細胞で合成され、肺胞腔へ分泌されます。これまでの研究で、SP-AやSP-Dは自然免疫の分野において、病原微生物に結合し増殖を抑制する、マクロファージの貪食を促進する、Toll様受容体を介した炎症を制御する、といった重要な役割を担っていることが明らかになっています。その一方で、SP-AやSP-Dが抗腫瘍作用も合わせ持つことを示唆する臨床的な報告がなされてきましたが、その分子メカニズムは明らかではありませんでした。
EGFRの細胞外ドメインにはN型糖鎖が存在し、EGFRの機能制御において重要であると考えられています。またガレクチン3やガングリオシドGM3といった糖鎖認識分子がEGFRの糖鎖を認識し、下流シグナルを制御するという報告がなされています (Partridge EA et al., Science 2004) (Yoon SJ et al., Proc Natl Acad Sci USA 2006) 。SP-AやSP-Dはグルコースやマンノースに対して高い親和性を有するレクチンであり、我々はSP-AやSP-DがEGFRの糖鎖へ作用する可能性について着目しました。
これまでの本学医化学講座との共同研究によって、SP-Dが肺がん細胞のEGFRの自己リン酸化と下流シグナルを抑制することで、肺がん細胞の増殖、遊走、浸潤を抑制することが明らかになりました。125I-EGFを用いた結合アッセイによって、SP-DはEGFとEGFRの結合を低下させることがわかりました。またEGFRの細胞外ドメインを精製し、SP-Dとの相互作用を解析した所、SP-DはEGFRのN型糖鎖に直接結合することがわかりました。さらに質量分析を用いて、EGFRのドメインⅢ(リガンド結合部位)に位置する328番目と337番目のアスパラギン残基に高マンノース型のN型糖鎖が存在することを確認しました。以上より、SP-DはEGFRの細胞外ドメインに存在する高マンノース型のN型糖鎖に糖鎖認識領域を介して結合し、EGFRのリガンド結合を阻害することにより、EGFシグナルを抑制し、抗腫瘍作用をもたらすと考えられました(Hasegawa Y et al., Oncogene 2014)。
SP-Aについても、肺がん細胞のEGFRに直接結合し、EGFシグナルを抑制し、細胞増殖・遊走・浸潤を抑制することがわかっています。現在、その詳細な分子メカニズムを解明すると共に、動物実験モデルにおける検討も開始しています。
肺サーファクタント蛋白質AとD(SP-A、SP-D)は肺胞Ⅱ型細胞やクララ細胞で合成され、肺胞腔へ分泌されます。これまでの研究で、SP-AやSP-Dは自然免疫の分野において、病原微生物に結合し増殖を抑制する、マクロファージの貪食を促進する、Toll様受容体を介した炎症を制御する、といった重要な役割を担っていることが明らかになっています。その一方で、SP-AやSP-Dが抗腫瘍作用も合わせ持つことを示唆する臨床的な報告がなされてきましたが、その分子メカニズムは明らかではありませんでした。
EGFRの細胞外ドメインにはN型糖鎖が存在し、EGFRの機能制御において重要であると考えられています。またガレクチン3やガングリオシドGM3といった糖鎖認識分子がEGFRの糖鎖を認識し、下流シグナルを制御するという報告がなされています (Partridge EA et al., Science 2004) (Yoon SJ et al., Proc Natl Acad Sci USA 2006) 。SP-AやSP-Dはグルコースやマンノースに対して高い親和性を有するレクチンであり、我々はSP-AやSP-DがEGFRの糖鎖へ作用する可能性について着目しました。
これまでの本学医化学講座との共同研究によって、SP-Dが肺がん細胞のEGFRの自己リン酸化と下流シグナルを抑制することで、肺がん細胞の増殖、遊走、浸潤を抑制することが明らかになりました。125I-EGFを用いた結合アッセイによって、SP-DはEGFとEGFRの結合を低下させることがわかりました。またEGFRの細胞外ドメインを精製し、SP-Dとの相互作用を解析した所、SP-DはEGFRのN型糖鎖に直接結合することがわかりました。さらに質量分析を用いて、EGFRのドメインⅢ(リガンド結合部位)に位置する328番目と337番目のアスパラギン残基に高マンノース型のN型糖鎖が存在することを確認しました。以上より、SP-DはEGFRの細胞外ドメインに存在する高マンノース型のN型糖鎖に糖鎖認識領域を介して結合し、EGFRのリガンド結合を阻害することにより、EGFシグナルを抑制し、抗腫瘍作用をもたらすと考えられました(Hasegawa Y et al., Oncogene 2014)。
SP-Aについても、肺がん細胞のEGFRに直接結合し、EGFシグナルを抑制し、細胞増殖・遊走・浸潤を抑制することがわかっています。現在、その詳細な分子メカニズムを解明すると共に、動物実験モデルにおける検討も開始しています。
肺がん幹細胞を誘導する分子メカニズムの解明
多くのがん研究によってがんの遺伝学的バックグラウンドが明らかになるにつれ、がんのモノクローナリティ(単一性)が強調されてきました。しかし1994 年のLapidot らによる急性骨髄性白血病(AML) 幹細胞の発見により、がん幹細胞理論ががん研究の主要なテーマとなりました。以後,固形腫瘍でもがん幹細胞分離の報告が続々となされ、肺がんに関しても世界中で研究が行われております。
がん幹細胞とは、(1)高い造腫瘍能を有し、(2)自己複製能を有し、(3)多分化能を有する細胞集団であり、非がん幹細胞と比較して、化学療法や放射線療法等の治療に対して抵抗性を示す事が明らかとなっております。つまり化学療法、放射線療法により、がんが一見縮小したかに見えても、実は非がん幹細胞だけが障害され、治療抵抗性を示すがん幹細胞は局所に残存している可能性があるという事を示唆します。さらには、治療により画像診断学的にがんを全て排除できたと考えられる場合でも、その後、ある一定の確率でがんを再発する臨床の実情をも示唆するものであります。これらの特徴を持つがん幹細胞を誘導する分子メカニズムの解明は、がん幹細胞を治療標的とした新たな治療法の開発に必要不可欠であると考えられます。
がん幹細胞を分離同定する方法はいくつか報告されておりますが、確定的な分子マーカーや、同定法が確立されている訳ではありません。
がん幹細胞様細胞としてこれまで報告された分離法としては、(1)細胞表面マーカーによる分離法、(2) side population 法、(3) ALDEFLUORアッセイ、(4)sphere formation assay などが知られております。現在我々も病理学第一講座と合同でこれらの方法を用い肺がん幹細胞様細胞を分離し、解析を行っております。
がん幹細胞とは、(1)高い造腫瘍能を有し、(2)自己複製能を有し、(3)多分化能を有する細胞集団であり、非がん幹細胞と比較して、化学療法や放射線療法等の治療に対して抵抗性を示す事が明らかとなっております。つまり化学療法、放射線療法により、がんが一見縮小したかに見えても、実は非がん幹細胞だけが障害され、治療抵抗性を示すがん幹細胞は局所に残存している可能性があるという事を示唆します。さらには、治療により画像診断学的にがんを全て排除できたと考えられる場合でも、その後、ある一定の確率でがんを再発する臨床の実情をも示唆するものであります。これらの特徴を持つがん幹細胞を誘導する分子メカニズムの解明は、がん幹細胞を治療標的とした新たな治療法の開発に必要不可欠であると考えられます。
がん幹細胞を分離同定する方法はいくつか報告されておりますが、確定的な分子マーカーや、同定法が確立されている訳ではありません。
がん幹細胞様細胞としてこれまで報告された分離法としては、(1)細胞表面マーカーによる分離法、(2) side population 法、(3) ALDEFLUORアッセイ、(4)sphere formation assay などが知られております。現在我々も病理学第一講座と合同でこれらの方法を用い肺がん幹細胞様細胞を分離し、解析を行っております。
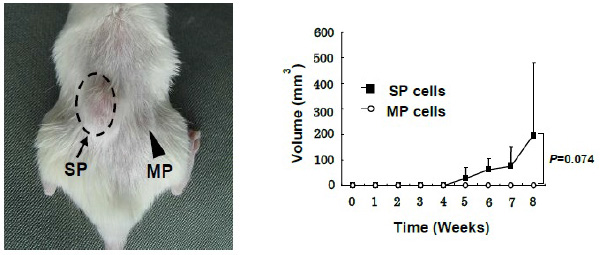
肺がん幹細胞様細胞の分離
ヒトの肺腺癌の細胞株であるLHK2を用い、side population 法によりがん幹細胞様細胞としてSP細胞、非がん幹細胞様細胞としてMP細胞を分離致しました。それらを免疫不全マウスに接種したところ、SP細胞は、MP細胞よりも有意差を持って高い造腫瘍能を持つことが証明されました。
肺がん幹細胞様細胞phenotypeにSOX2発現が関与する
以前我々は、LHK2のSP細胞においてSOX2の発現が高いことをRT-PCRにて証明致しました。また、SOX2過剰発現細胞はSP細胞の割合が増加し、コントロール細胞に比べ高い造腫瘍能を持ち、またSOX2 knockdown細胞はコントロール細胞よりも低い造腫瘍能を持つことを証明致しました。つまりSOX2の発現は肺がん幹細胞様細胞phenotypeに関与することがわかりました。
現在我々は、肺がん幹細胞様細胞のSOX2を誘導する分子メカニズムの解明を目的に研究を進めております。
現在我々は、肺がん幹細胞様細胞のSOX2を誘導する分子メカニズムの解明を目的に研究を進めております。
免疫エスケープ腫瘍を標的とする新しいペプチドワクチン開発
CD8+T細胞(CTL)は細胞障害活性を持っており、ウイルスなどの細胞内病原体に感染した細胞を傷害します。ペプチドワクチン療法はCTLの細胞障害活性を利用した治療で、癌に特異的にみられる抗原ペプチド(癌ペプチド)を投与することで標的細胞(癌細胞)に特異的なCD8+T細胞を活性化、増殖させ癌細胞を攻撃させる治療です。CD8+T細胞は、癌細胞のMHCクラスI分子に提示される抗原ペプチドを認識しますが、通常これらのペプチドは抗原プロセシングと呼ばれる複雑かつ精緻にコントロールされた過程を経て細胞内でつくられています。
このプロセシング過程の中で、MHCクラスI分子はPLC(peptide loading complex)と呼ばれる複合体を形成することが知られており、これはMHC classIと結合する安定した抗原ペプチドの結合の選択に必要であると考えられています。私はPLCの構成分子の一つであるtapasinというシャペロンタンパクに注目しました。その理由は、多くの癌組織においてtapasinの発現低下が認められるからです。Tapasinの発現が低下するとPLCの形成が出来なくなり、その結果、細胞表面に提示されるMHCクラスI ペプチドは減少しCTL認識からのエスケープが可能になります。これはこれからのペプチドワクチン治療を考えるうえで大きな問題となります。私たちは病理学第一講座と合同でtapasin遺伝子を欠損したマウスモデルを利用し、このような免疫エスケープ腫瘍を標的可能とする、これまでにない新しいタイプのペプチドワクチン開発に携わっています。
このプロセシング過程の中で、MHCクラスI分子はPLC(peptide loading complex)と呼ばれる複合体を形成することが知られており、これはMHC classIと結合する安定した抗原ペプチドの結合の選択に必要であると考えられています。私はPLCの構成分子の一つであるtapasinというシャペロンタンパクに注目しました。その理由は、多くの癌組織においてtapasinの発現低下が認められるからです。Tapasinの発現が低下するとPLCの形成が出来なくなり、その結果、細胞表面に提示されるMHCクラスI ペプチドは減少しCTL認識からのエスケープが可能になります。これはこれからのペプチドワクチン治療を考えるうえで大きな問題となります。私たちは病理学第一講座と合同でtapasin遺伝子を欠損したマウスモデルを利用し、このような免疫エスケープ腫瘍を標的可能とする、これまでにない新しいタイプのペプチドワクチン開発に携わっています。
EGFR遺伝子変異、ALK融合遺伝子とサーファクタント蛋白質の関連
肺がんの一部にはEGFR遺伝子変異やALK融合遺伝子など、特有な遺伝子異常が存在することが明らかとなっています。この様な遺伝子異常をターゲットにした分子標的治療薬が使用可能となり、肺がん治療は大きく進歩しました。肺サーファクタント蛋白質のひとつであるSP-Aは肺に特異的に発現することから、肺腺がんの組織診断マーカーとして広く使用されていますが、肺サーファクタント蛋白質の肺がんにおける発現については、未だその詳細はよくわかっていません。わたしたちは肺サーファクタント蛋白質の肺がんにおける発現について、特にEGFR遺伝子変異やALK融合遺伝子との関連について調査し、肺癌の発生病理について検討しています。さらに血清SP-A、SP-D値と肺がん治療や予後などとの関連について検討を行い、肺がん診療における肺サーファクタント蛋白質測定の有用性について検討を行っています。
最終更新日:2015年03月18日